Genetik und Epigenetik
________________________________________________________________
Genetik
In der Genetik oder Vererbungslehre geht es darum, die Gesetzmäßigkeiten und die materiellen Grundlagen, die zur Ausprägung von erblichen Merkmalen führen, zu erforschen.
Mendelsche Regeln
Gregor Mendel publizierte seine Vererbungsregeln in 1866. Er
stellte 3 Regeln auf, die er durch Beobachtung heraus fand:
1. Uniformitätsregel
2. Spaltungsregel
3. Unabhängigkeits/Neukombinationsregel
In den Körperzellen diploider Organismen treten die Chromosomen
paarweise auf. So besitzt der Hund 39 Chromosomenpaare, also 78
Chromosomen. Je eines der Chromosomen in einem Chromosomenpaar stammt vom
Rüden und eines von der Hündin. Die beiden Chromosomen eines Paares werden
auch als homologe Chromosomen bezeichnet. In einer Körperzelle sind die
Erbanlagen pro Merkmal somit immer doppelt vorhanden.
Bei der Bildung der Geschlechtszellen werden die homologen
Chromosomenpaare in der Meiose getrennt. In einer Eizelle bzw.
einem Samenfaden befindet sich also nur der einfache Chromosomensatz, die
Erbanlagen pro Merkmal (Genabschnitte) sind somit immer nur einmal
vorhanden. Dies erklärt die Spaltungsregel.
Bei der Befruchtung, also der Verschmelzung von Eizelle und
Samenfaden, bringen beide Geschlechtszellen jeweils eine Erbanlage pro
Merkmal mit. Die durch diese Verschmelzung entstehende befruchtete
Eizelle (Zygote) hat also wieder den doppelten Chromosomensatz. Die
Erbanlagen können so neu kombiniert werden. Dies erklärt die
Unabhängigkeitsregel.

__________________________________
Grundlagen angewandter Genetik für die Hundezucht
Um Erbkrankheiten bekämpfen zu können, muss man sich mit Genetik
auskennen. Man sollte wissen, wie Erbgänge funktionieren, wie man ungefähr
die Anzahl der latent betroffenen Hunde schätzen kann und welche Maßnahmen
möglich und im Einzelfall angezeigt sind. Aufgrund zahlreicher Anfragen
habe ich versucht, den Kurzvortrag, den ich auf der JHV 2010 hielt, in
Artikelform umzuarbeiten.
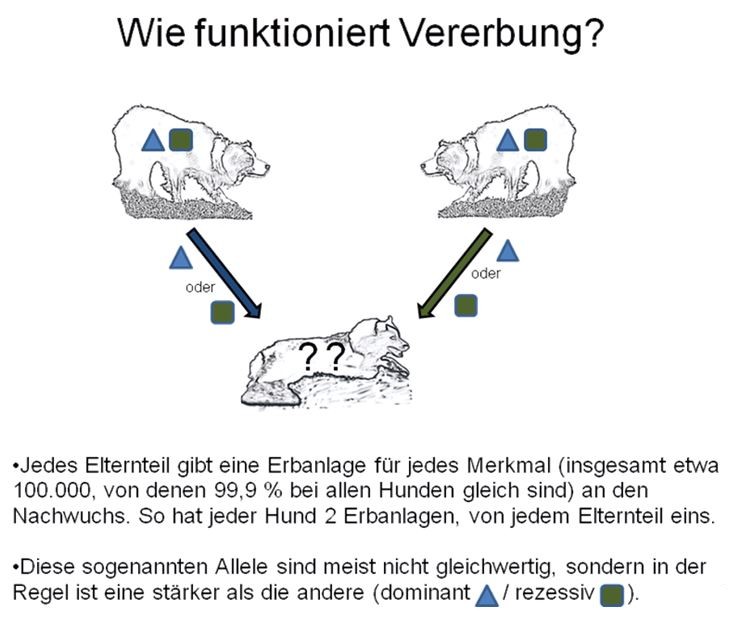
Die Erbanlagen bewirken die Ausbildung aller körpereigenen Merkmale, sie
sind quasi der Bauplan des Körpers. Starke Genanlagen können bereits in
einfacher Kopie als Bauplan dienen, schwache Genanlagen müssen in
doppelter Kopie vorliegen, damit sie ausgeführt werden. Diese starken
Anlagen nennt man dominant, die schwachen heißen rezessiv.
Hunde, die
zwei gleiche Erbanlagen haben, nennt man reinerbig oder homozygot. Hunde,
die unterschiedliche Erbanlagen haben, nennt man mischerbig oder
heterozygot.
Erbkrankheiten, die über dominante Erbanlagen vererbt
werden, sind einfach zu bekämpfen. Ein einzelnes Allel reicht aus, um sie
auszubilden. Daher kann man auch die mischerbig Betroffenen leicht
erkennen und aus der Zucht nehmen.
Problematisch sind immer die rezessiven Erkrankungen. Zum Ausbruch dieser
Erkrankungen muss die doppelte Kopie des rezessiven Gens vorliegen. Die
mischerbig Betroffenen sind immer selbst gesund. Sie sind von den
reinerbig Gesunden optisch nicht zu unterscheiden. Mischerbige Träger von
rezessiven Merkmalen nennt man Carrier. Die modernste und beste
Möglichkeit, Carrier zu erkennen, ist über einen Gentest. Leider gibt es
Gentests nur für wenige Erkrankungen wie z.B. CEA. Am Beispiel der CEA
haben wir gesehen, dass Carrier völlig harmlos sind, solange man sie an
reinerbig Gesunde anpaart. Ein einzelnes, krankmachendes rezessives Gen
kann sich gegen eine gesunde dominante Kopie nie durchsetzen.
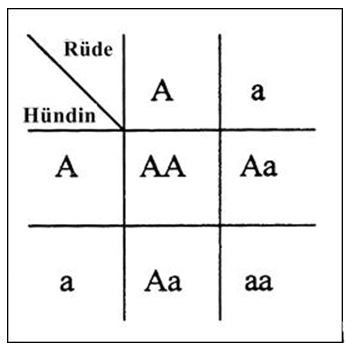
Die folgende kleine Tabelle zeigt, welche Genkombinationen möglich sind,
wenn 2 Carrier aneinander angepaart werden. Das dominante Gen wird mit
Großbuchstaben gekennzeichnet, das rezessive Gen mit kleinem Buchstaben.
Bei der Häufigkeitsverteilung fällt auf, dass doppelt so viele Carrier
entstehen werden wie reinerbige Nachkommen. Solch eine Verteilung würde
man beispielsweise bei CEA erwarten.
Nun sind aber leider nur die wenigsten Merkmale durch ein einzelnes Gen
bestimmt. Meist sind mehrere Gene beteiligt. Einen solchen Erbgang nennt
man polygenen Erbgang. HD ist ein Beispiel für polygene Vererbung. Auch
für Epilepsie steht im Raum, dass es sich um einen polygenen Erbgang
handelt.
Wie viele und welche Möglichkeiten gäbe es nun bei einem Merkmal, das von
3 Genen beeinflusst wird?
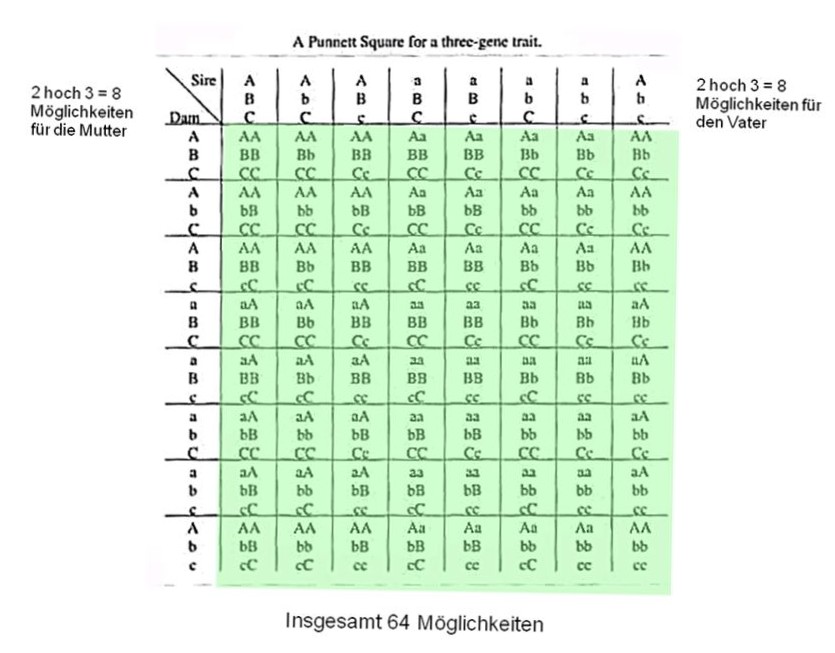
Bei 8 mal 8 Ausgangspositionen kommt man auf insgesamt 64 verschiedene
Genverteilungen, die bei den Nachkommen auftreten können.
Wie sähe die zahlenmäßige Verteilung der verschiedenen Varianten aus?
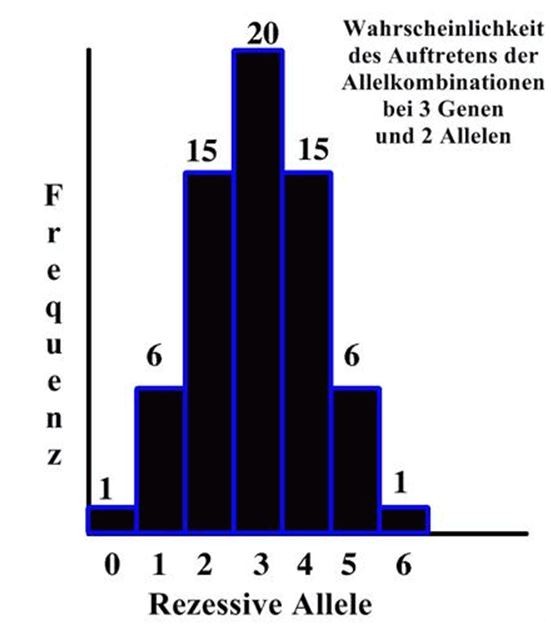
Die statistische Analyse der theoretischen Möglichkeiten ergibt lediglich
einen einzigen Nachkommen, der reinerbig dominant, also völlig frei von
möglichen Krankheitsanlagen wäre. Dann kommen 6 Hunde mit einem rezessivem
Gen und 5 dominanten, 15 Hunde mit 2 rezessiven und 4 dominanten, 20 Hunde
mit jeweils 3 rezessiven und 3 dominanten Genen, wieder 15 Hunde mit 4
rezessiven und 2 dominanten Genen und schlussendlich ein einziger Hund,
der 6 rezessiven Gene abbekommen hat und daher krank wird.
Wichtig daran ist, dass es umso mehr Carrier gibt, je mehr Gene an der
Ausprägung eines Merkmales oder einer Erkrankung beteiligt sind.
Einer
relativ kleinen Anzahl reinerbig Kranker und reinerbig Gesunder, nämlich
jeweils nur einem, steht eine große Anzahl Mischerbiger gegenüber. Von je
mehr Genen eine Erkrankung verursacht wird, umso mehr Carrier kann man
statistisch erwarten. Uns beschäftigen in der Regel die polygen vererbten
Merkmale.
Wie funktioniert eine polygene Vererbung?
Für Verwirrung sorgt die Tatsache, dass es verschiedene Bezeichnungen für
Erbgänge gibt, bei denen verschiedene Gene zusammen- oder gegeneinander
wirken. So findet man die Bezeichnungen polygen rezessive Vererbung,
polygen additive Vererbung ebenso wie multifaktorielle Vererbung. Alle
bezeichnen mehr oder weniger dasselbe.
Viele Krankheiten und Merkmale zeigen eine familiäre Häufung, ohne dass
die Zahlen einem klassischen, auf einem Genpaar beruhenden (monogen)
Erbgang entsprechen würden. Verwandte ersten Grades (Eltern, Kinder,
Vollgeschwister) sind zumeist in etwa 2-4% betroffen und damit wesentlich
geringer als aufgrund einer monogenen Vererbung. Die gängige Hypothese
ist, dass hierbei mehrere Gene additiv und unabhängig voneinander sowie
die verschiedensten Umweltfaktoren zusammenwirken. Meist wird dieses
Modell als „multifaktorielle Vererbung“ bezeichnet. Damit wird schon im
Namen klar, dass viele Faktoren zusammenkommen müssen, um ein bestimmtes
Merkmal auszuprägen, bzw. eine Krankheit auszulösen. Da es sich häufig um
Merkmale handelt, die fließend von sehr gering bis sehr ausgeprägt
vorhanden sein können, spricht man auch von einer quantitativen Vererbung.
In diesem Modell wirken die verschiedenen Gene und Umweltfaktoren und
erzeugen eine Anfälligkeit (Prädisposition). Beispiele für derartige,
nicht krankhafte Merkmale sind Körperhöhe und –statur, aber auch
Verhaltensanlagen wie Intelligenz beim Menschen oder Arbeitsveranlagung
bei unseren Hunden.
Den Anteil der Erbanlagen an der Ausprägung eines Merkmales (einer
Erkrankung) nennt man Heritabilität. Heritabilität und Umwelteinflüsse
ergeben logischerweise zusammen immer das Ganze, was mit 1 oder 100%
bezeichnet wird. Bei einem Heritabilitätskoeffizienten von 0,4 (40%) wird
also knapp die Hälfte der auslösenden Faktoren von den Genen bestimmt und
60% von der Umwelt. Je weniger unterschiedlich die Umwelteinflüsse
innerhalb einer Population sind, umso stärker kommt die ererbte
„Grundausstattung“ zum Tragen. Ein Beispiel: bei normaler Ernährung wird
die Größe überwiegend von den Erbanlagen bestimmt. Lässt man Welpen jedoch
fast verhungern, bleiben sie lebenslang kleiner als Geschwister, die
normal ernährt werden.
Die Anlagen für multifaktoriell vererbte Krankheiten sind zwar innerhalb
der Population (hier: Hunde in einem Zuchtbuch) auch fließend vorhanden,
die Krankheit selbst tritt aber meist entweder auf oder nicht. Aus diesem
Grund wurde der sogenannte Schwellenwert eingeführt. Der Schwellenwert
gibt an, ab welcher Anzahl von Genen und Umweltfaktoren eine Erkrankung
auftritt.
Funktionsweise einer polygenen Vererbung am Beispiel HD:
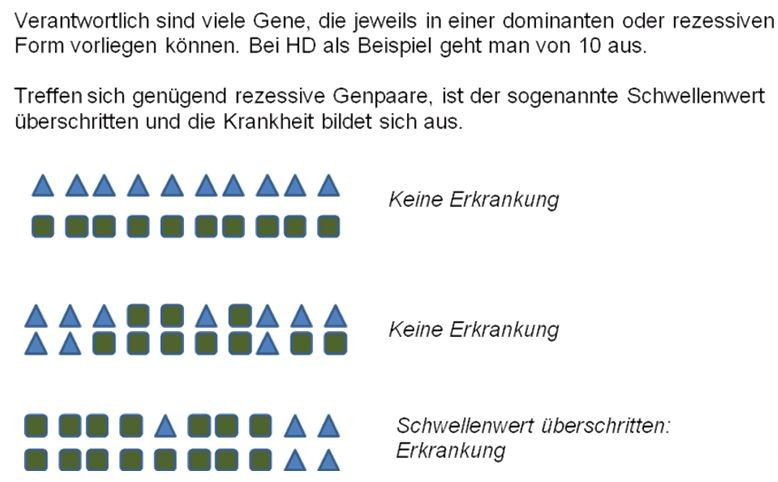
In der ersten Zeile sehen wir die fiktive Genanordnung eines Hundes, der
von einem Elternteil ausschließlich dominante (Dreieck) und vom anderen
ausschließlich rezessive Gene (Quadrat) mitbekommen hat. Dieser Hund wäre
gesund, weil die dominanten Gene ja in jedem einzelnen Fall stärker als
die rezessiven Kopien wären. Für die Zucht könnte dieser Hund aufgrund
seiner hohen Zahl an rezessiven Genen aber selbstverständlich
problematisch sein. Dies wäre beispielsweise eine Konstellation eines
Hundes mit exzellenten Hüftgelenken, der aber HD vererbt. Das erklärt
auch, warum es immer sicherer ist, auf die ganze Familienleistung zu sehen
als auf den einzelnen Hund. In der Familie dieses Hundes wird es etliche
Hunde mit schlechteren HD-Ergebnissen geben.
Die mittlere Zeile zeigt
drei reinerbig rezessive Genpaare, einige gemischte und einige reinerbig
dominante. Die drei rezessiven Genpaare sind zu wenig, um den
Schwellenwert zu erreichen, der in unserem Beispiel bei 6 Genpaaren liegt.
Der letzte Hund hat 7 rezessive Paarungen abbekommen. Er wird krank
werden, denn der Schwellenwert ist überschritten.
… und wie sieht die Verteilung der einzelnen Genkombinationen bei einer
polygenen Vererbung aus?
Bei 10 Genpaaren gibt es 2 hoch 10 Verteilungsmöglichkeiten für jedes
Elternteil. Das sind 1064 pro Elternteil. Für Nachkommen gibt es dann 1024
x 1024, also über eine Million verschiedende Möglichkeiten von
Allelkombinationen. Das kann man nicht mehr in Stufen anzeigen, sondern es
ergibt eine Kurve.
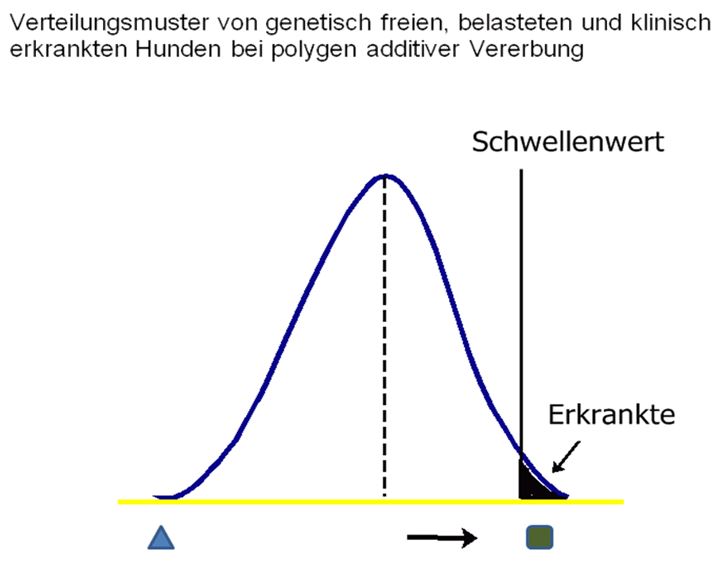
Ähnlich der Stufengrafik weiter oben ist auch hier die Anzahl der völlig
freien (rein dominanten) ganz links zu finden. Dann kommt die große Menge
der Mischerbigen und dann die wieder sinkende Anzahl der Hunde mit
überwiegend rezessiven Genen. Jenseits des Schwellenwertes liegen die
Hunde, die so viele rezessive Paare haben, dass die Krankheit bei ihnen
ausbricht.
In einer betroffenen Familie zeigt das Auftreten der Erkrankung, dass mehr
Gene vorhanden sind, so dass die Normalkurve nach rechts verschoben wird.
Die Inzidenz der Erkrankung und damit auch das Wiederholungsrisiko in der
Familie wird größer.
Das Wichtigste an dieser Kurve ist, dass sie einem zeigt, wie viele
Carrier es im Vergleich zu den erkrankten Hunden gibt, und dass es einem
Hund nicht anzusehen ist, wo auf der Kurve er sich befindet. Ein enger
Verwandter eines Erkrankten hat eine höhere Wahrscheinlichkeit, auf der
rechten Seite der Kurve zu liegen – er kann aber auch ganz links liegen.
Wo er sich tatsächlich befindet, wird erst über seine Nachkommen sichtbar.
Die Wahrscheinlichkeit, dass hohe rezessive Genfrequenzen vorliegen, sind
ganz allgemein bei Verwandten ersten Grades (Eltern, Kinder, Geschwister)
eines Erkrankten am höchsten. Das Wiederholungsrisiko entspricht der
Quadratwurzel der Inzidenz. Bei einer angenommenen Inzidenz von 5% kommt
man zu einem Wiederholungsrisiko von etwa 22 Prozent.
Entferntere
Verwandte haben ein viel kleineres Wiederholungsrisiko, da sie über ihre
anderen Verwandten auch ganz anderes Genmaterial besitzen. Bei
Halbgeschwistern oder Kindern von Vollgeschwistern, beispielsweise, sinkt
das Risiko schon auf die Hälfte desjenigen von Vollgeschwistern. Die beste
Einschätzung über den Genstatus eines einzelnen Hundes bekommt man über
die Ergebnisse von Nachkommen.
Möchte man Erbkrankheiten vermeiden und kennt die aufgetretenen Fälle in
den Pedigrees der potentiellen Zuchtpartner, lässt sich das Risiko für
jede geplante Paarung errechnen.
Wieso kann man nicht einfach die Carrier ausmerzen?
Mindestens 40% aller Hunde, die wir züchten, haben irgend einen Defekt,
und sei er auch noch so klein. Im Durchschnitt trägt jeder Hund rezessive
Anlagen für 5 Defekte. Es gibt also massenhaft Carrier für irgendeinen
Defekt. Wollte man die alle aus der Zucht nehmen, bliebe fast nichts
übrig.
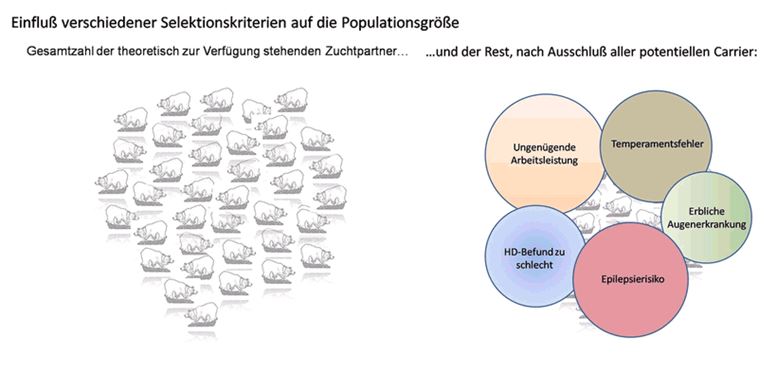
Hunderassen sind künstlich geschlossene Populationen. Border Collies
wurden in der ISDS seit 100 Jahren in einem geschlossenen Zuchtbuch
gezüchtet. Das bedeutet, dass sich zwar durch Vermehrung die Gesamtzahl
der Hunde enorm vergrößert hat, es ist aber so gut wie kein neues
Genmaterial mehr hinzugekommen. Es gibt zwar inzwischen Tausende von
Border Collies, aber diese sind untereinander verwandter als in früheren
Tagen. Das sieht man natürlich nur, wenn man die Stammbäume über
Jahrzehnte verfolgt.
Mit steigendem Verwandtschaftsgrad steigt auch das
Risiko, dass sich gleiche Genpaare zusammenfinden, die Population wird
reinerbiger – und das Risko, dass sich rezessive Defektgene
zusammenfinden, steigt auch.
Diese sogenannte Verkleinerung des
Genpools ist eine langsam schleichende Entwicklung. Sie wird von vielen
Fachleuten als das Hauptproblem der Hundezucht in der Zukunft gesehen. Das
immer häufigere Auftreten von „neuen“ Erbkrankheiten hat nicht nur mit den
verbesserten Diagnosemöglichkeiten der Tiermedizin in den letzten
Jahrzehnten zu tun, sondern es ist tatsächliche Folge des steigenden
Verwandtschaftsgrades.
Bezüglich der Bekämpfung von Erbkrankheiten ergeben sich folgende
Schlüsse: Eine züchterische Merzung (Ausschluss aus der Zucht) für
Merkmalsträger und mögliche Carrier ist erfolgversprechend bei einer
Inzidenz von <1%.
Die Inzidenz von beispielsweise Epilepsie beim Hund
(alle Rassen) ist etwa 5%. Die Carrierfrequenz bei einer Inzidenz von 5%
und angenommener polygener Vererbung liegt, grob geschätzt, bei 40 %!
Nimmt man signifikante Anzahlen von Hunden aus der Zucht, so bedingt dies
ganz zwangsläufig das man das Auftreten neuer Erbkrankheiten fördert. Für
jeden Kopf der Hydra, den man abschlägt, wachsen neue nach.
Darüber
hinaus kann die Entfernung von Hunden aus dem Genpool noch weitere
Konsequenzen haben, die man gar nicht beabsichtigt hat.
Erstens ist es
keineswegs so, dass krankmachende Gene nur isoliert zu betrachten wären.
Es kann durchaus sein, dass in direkter Nachbarschaft auf einem Chromosom
Gene für sehr wichtige Eigenschaften liegen. Diese würde man gleich mit
ausmerzen, ohne es zu wollen.
Zweitens ist eine Krankheit manchmal nur
das aus dem Gleichgewicht geratene Extrem eines an sich sehr notwendigen
und wertvollen Stoffwechselweges. Betrachten wir die Kurve mit dem
Schwellenwert, so ist es häufig so, dass die fürs Überleben beste
Genvariante auf der Spitze der Kurve liegt. Dann wäre es ganz unproduktiv,
danach zu streben, nur noch die reinerbig Dominanten übrigzubehalten, denn
sie sind genau so weit weg vom Ideal wie die Kranken.
Bei Epilepsie, beispielsweise, nimmt man an, dass der Transport von
Natrium und Calcium durch die Nervenzellwand, also die konkrete chemische
Reaktion zur Erzeugung von Potentialdifferenzen und damit der
Nervenleitfähigkeit, gestört ist. Der Transport an sich ist aber notwendig
für jeglichen Denk- oder Bewegungsvorgang. Es kann durchaus sein, dass
Hunde mit Epilepsie ein aus den Fugen geratenes Transportsystem haben, die
mischerbigen Verwandten aber ein besonders schnelles oder besonders
leistungsfähiges Transportsystem haben.
Das weiß niemand, aber sogar
Molekulargenetiker, die Gentests entwickeln, warnen davor ihre Tests zum
Zuchtausschluss ganzer Gruppen zu verwenden. Die renommierte Genforscherin
Vicki Meyers-Wallen schreibt dazu: „Solange wir nichts über die
Interaktionen zwischen Genen wissen, wissen wir nicht, ob nicht einige
dieser Gene andere Funktionen haben, die wir behalten möchten. Auch andere
populationsgenetische Effekte sollten nicht ignoriert werden. Es könnte
sein, dass es am besten ist, diese Informationen nur dazu zu benutzen, um
Paarungen zu vermeiden, aus denen Kranke entstehen können anstatt ganze
Gengruppen aus der Population zu entfernen und eine möglichst große
Genvielfalt zu erhalten. Daher sollten wir Züchter nicht dazu beraten, bei
polygenen Vererbungen ganze Gruppen zu selektieren, ehe wir nicht mehr
über Interaktionen zwischen Genen wissen. Es gibt nicht nur die
Möglichkeit, die Rassegesundheit über Selektionen zu verbessern sondern
auch die Gefahr, Schaden anzurichten, wenn wir es versäumen, die
Genvielfalt zu erhalten.
Diese eher theoretischen Aussagen bestätigen die alte Züchterweisheit,
dass die Mischung von Genen häufig die besten Ergebnisse erzielt. Das
klassische Beispiel ist der sogenannte Heterosiseffekt. Schwierig wird es,
wenn man diese Effekte stabilisieren will, denn das wiederum geht nur
durch Erhöhung der Reinerbigkeit. Erfolgreiche Zucht ist immer eine
Gradwanderung zwischen Outcross und Linienzucht. Beim Auftreten von
Erbkrankheiten ist das Pendel zu weit zur Linienzucht ausgeschlagen, und
es wird dringend Zeit zu mehr Auskreuzung.
Was tun?
Die Empfehlungen von Populationsgenetikern gehen alle dahin, Paarungen zu
vermeiden, bei denen das Risiko sehr hoch ist, dass sich rezessive
Genpaare treffen, und statt dessen „gefährdete“ Partner immer an möglichst
„ungefährdete“ anzupaaren. Das nennt man Vergrößerung der Heterogenität,
und es bedeutet, dass man die Anzahl der Mischerbigen (Heterozygoten)
erhöht. Damit kann das Auftreten von rezessiven Erbkrankheiten vermieden
werden. Wir erinnern uns, eine rezessive Kopie kommt niemals gegen eine
dominante Kopie an, der konkrete Bauplan des Hundes wird vom dominanten,
in diesem Fall nicht krankmachenden Gen, bestimmt. Darüberhinaus hat
diese Vorgehensweise den positiven Begleiteffekt, dass züchterisch
wertvolle Tiere, die ein erhöhtes Vererbungsrisiko für Krankheit tragen,
in der Zucht bleiben können.
Die komplette Ausmerzung von Erbkrankheiten ist leider reines
Wunschdenken. Eine populationsgenetische Untersuchung der Tierärztlichen
Hochschule Hannover zur „Zuchtsituation und erblich determinierten
Erkrankungen bei Golden und Labrador Retrievern“ kam zu der ernüchternden
Feststellung, dass „eine absolute Selektion gegen all diese Defekte
gleichzeitig nicht möglich ist (…) und ein Selektionsfortschritt nur sehr
langsam und nur bei Indexselektion zu erwarten wäre. (…) Die einzige
Möglichkeit, diese Erkrankungen in ihrer Ausprägung schneller einzudämmen,
ist eine Zucht auf größere Heterogenität innerhalb der Populationen.“
Ähnlich äußern sich andere Populationsgenetiker, wie zum Beispiel, Dr.
Irene Sommerfeld-Stur aus Wien. „Der Erhalt der genetischen Varianz ist
eines der wichtigsten Ziele der modernen Hundezucht. (…) Fatalerweise
führen oft gerade züchterische Entscheidungen, die im Interesse der
Gesundheit der Nachkommen liegen, zu einer Verschärfung der Situation der
genetischen Vielfalt. (…) Es entspricht also nahezu der Quadratur des
Kreises die Forderung nach Erhalt der genetischen Vielfalt mit den
ebenfalls vielfältigen Selektionszuchtzielen unter einen Hut zu bringen.
Die einzige Möglichkeit, dieses Problem halbwegs erfolgreich zu lösen
bietet die Erarbeitung rassespezifischer Zuchtstrategien, die die
individuelle Situation der einzelnen Rassezuchtpopulationen
berücksichtigen und dabei alle Möglichkeiten der modernen Genetik
ausschöpfen. (…)
Es gibt eine Reihe von züchterischen Instrumenten, die in diesem
Zusammenhang genutzt werden können.
-
Berücksichtigung des
Inzuchtkoeffizienten oder des Ahnenverlustkoeffizienten
-
Zuchtwertschätzung
-
Selektion nach einem
Selektionsindex
-
Einsatz
molekulargenetischer Diagnoseverfahren“
Das hört sich alles sehr kompliziert an. Was versteht man unter diesen
Maßnahmen?
Theoretisch findet man in einem Pedigree zwei Eltern, 4 Großeltern, 8
Urgroßeltern und so weiter. Das macht bis zur dritten Generation 14
verschiedene Vorfahren. Der Ahnenverlustkoeffizient bezeichnet die Anzahl
von Vorfahren, die in einem Pedigree fehlen, weil sie mehrfach vorhanden
sind. Ist der Großvater mütterlicherseits gleichzeitig der Urgroßvater
väterlicherseits, sind es nur noch 13 Vorfahren. Mit dem
Ahnenverlustkoeffizient kann der Inzuchtgrad bestimmt werden.
Der
Inzuchtkoeffizient macht dasselbe, ist aber genauer und wird weiter nach
hinten berechnet als der meist von Hand ausgezählte
Ahnenverlustkoeffizient.
Ein hoher Ahnenverlust bedeutet also einen
hohen Verwandtschaftsgrad und damit eine höhere Wahrscheinlichkeit, dass
sich rezessive Gene treffen. Das kann gut sein, wenn auf Leistungsträger
Linie gezüchtet wird, wenn es sich herausstellt, dass der gemeinsame
Verwandte eine Krankheit vererbt, geht der Schuss nach hinten los.
Eine Zuchtwert- und damit Risikoeinschätzung für eine geplante Paarung
kann ein Züchter grob selbst bestimmen, wenn er weiß, welche
Erbkrankheiten in welchen Linien verlaufen und wie die Eigenleistung sowie
die Verwandtschaftsgrade zu Erkrankten beim potentiellen Zuchtpartner
sind.
Bei der genauen, computergestützten Version wird der Zuchtwert
eines Tieres über seine eigenen Eigenschaften und über die Eigenschaften
seiner Verwandten (Nachkommen, Eltern, Geschwister) berechnet. Dazu wird
ein rechnerischer Mittelwert des durchschnittlichen Zuchtwertes für ein
bestimmtes Merkmal erstellt. Dieser Mittelwert wird der Einfachheit halber
meist mit 100 bezeichnet, darunter liegende Werte sind besser als der
Durchschnitt, darüber liegende schlechter. Mit jeder Information über
einen Hund verändert sich nun dessen individueller Wert. Hunde ohne kranke
Verwandte haben einen niedrigeren Wert als 100. Hat ein Hund kranke
Verwandte, steigt sein Zuchtwert (das Risiko), je nach Anzahl und
Verwandtschaftsgrad über den Mittelwert. Hat solch ein Hund
Nachkommen, die nicht krank werden, sinkt sein Wert wieder, denn das ist
ein Hinweis darauf, dass er selbst trotz seiner Verwandtschaft nur wenige
krankmachende Gene mitbekommen hat. Jede neue Information führt dazu, dass
der Wert aller verwandten Hunde neu berechnet wird. Das macht dieses
System sehr präzise und aktuell. Wenn man mit einem Hund züchten will, so
muss man eine Zuchtwertschätzung beantragen. Liegt der Wert über 100 hat,
so ist die Zucht nur möglich, wenn man einen Zuchtpartner aussucht, der
einen entsprechend niedrigen Wert besitzt. Hier kann jeder Zuchtverband
eigene Grenzen setzen. Die Kosten sind gering, wenn die Abstammungen und
Grundinformationen einmal eingepflegt sind. Dann müssen nur noch neue
Daten eingegeben werden. Verschiedene Zuchtverbände finanzieren eine
Zuchtwertschätzung über einen kleinen Solidarbeitrag, der zusätzlich zum
Deckschein oder dem Wurfbogen bezahlt wird.
Bei der Indexselektion wird die Selektion für verschiedene Merkmale
gewichtet. Der Index bestimmt die Wertigkeit der verschiedenen
Selektionskriterien. Überragende Werte bei einem Merkmal können Schwächen
bei einem anderen Merkmal ausgleichen, um die Genvielfalt zu erhalten.
In der Nutztierzucht werden zur Verbesserung der Erbeigenschaften einer
Rasse schon seit langer Zeit Zuchtwertschätzung und Indexselektion
eingesetzt. Sie sind Mittel der Wahl bei polygener Vererbung und haben in
den letzten 15 Jahren auch ihren Weg in die Hundezucht gefunden. Das
erfahrenste, deutsche Institut in punkto Zuchtwertschätzung bei Hunden ist
das Rechenzentrum für Tierzucht und angewandte Genetik der Universität
Gießen. Etliche Hundezuchtvereine sind hier Kunden, anfänglich alle wegen
HD, später wegen aller möglichen Erbkrankheiten.
Nachbemerkung zur praktischen Anwendung:
Egal, für welche Methode man sich entscheidet, der Erfolg steht und fällt
in jedem Fall mit der Zuverlässigkeit der zugrundeliegenden Information.
Fehlende Informationen über Erkrankte führen dazu, dass Züchter bei der
Zuchtplanung in falsche Sicherheit gewiegt werden. Zum Beispiel Epilepsie
lässt sich sagen, dass die wenigen, wirklich verheerenden Würfe bei
unseren Border Collies sicher auch hätten vermieden werden können, wenn
die Züchter eine Chance gehabt hätten, das Risiko der geplanten Anpaarung
zu erkennen. Niemand züchtet vorsätzlich kranke Tiere, schon gar nicht bei
dieser schrecklichen Krankheit.
Solange Züchter oder Deckrüdenbesitzer
Informationen über erkrankte Hunde zurückhalten, weil sie befürchten, dass
dann auch niemand mehr ihre Welpen aus anderen Würfen kauft oder dass auch
ihre anderen Hunde von der Zucht ausgeschlossen werden, solange wird eine
effektive Bekämpfung ein Wunschtraum bleiben.
Auch in der momentanen
Situation zeigt sich, dass sogar Züchter, die laut für die Bekämpfung von
Epilepsie werben, ihre eigenen Fälle keineswegs publik machen. Stattdessen
sind die, die ehrlich mit gutem Beispiel voran gegangen sind, die
einzigen, über die geredet wird. Das ist weder fair noch sinnvoll,
überlässt es doch wieder nur dem Zufall das Feld.
Was wir brauchen, ist eine Lösung für die Züchter, die Spielraum für gute
Arbeitshundezucht lässt und gleichzeitig das Risiko minimiert und damit
die Welpenkäufer schützt. Verwendet man das zugängliche Wissen der
Spezialisten auf diesem Gebiet, der Genetiker, ist das ist relativ einfach
durch kluge Anpaarung machbar, die Karten müssen aber endlich auf den
Tisch.
Dr. Viola Hebeler
Literaturverzeichnis folgt.